
NEWS
点击蓝字 关注我们
对于有性繁殖的动物来说,选择最佳的配偶对于最大化繁殖适应性非常重要。以秀丽隐杆线虫为例,其种群繁殖主要为雌雄同体自体受精,但也存在雄性与雌雄同体交配受精。在此背景下,雄性线虫会识别并偏好特定雌雄同体作为配偶,这种识别能力建立在一系列精细的感官评估之上,涉及对多种环境及生物信号的检测与综合处理。已知的是,雌雄同体的发育阶段(雄虫对L4及以上雌雄同体有偏好,对L3时期线虫则缺乏兴趣)构成了影响雄性偏好的一个显著因素,那么还有哪些因素会影响雄性对配偶的偏好呢?从今年3月份发表的“C. elegans males optimize mate-preference decisions via sex-specific responses to multimodal sensory cues”这篇文章中,我们可以获取许多有趣的信息。
影响因素一:天门冬苷信息
既往研究已经确定了雌雄同体来源的多种分泌和表面附着线索,这些线索在吸引及维系雄性伴侣方面展现出显著效能。其中最典型的特征是天门冬苷信息素,如主要由雌雄同体产生的ascr#3和ascr#8。这些化合物对雄性滞留具有强效应,并且根据环境的不同,能够排斥雌雄同体,从而调节种群内的社会互动。
那么,这种无挥发性的内源性天门冬苷对雄性配偶偏好决定的贡献到底有多大呢?
daf-22是合成短链天门冬苷(包括所有已知的天门冬苷信息素)所必需的酶,而daf-22突变虫株不能合成天门冬苷。实验发现(方法见图1),雄性线虫很容易识别daf-22突变成虫的性别,雄性更喜欢与daf-22突变的雌雄同体发生相互作用,而非daf-22突变雄虫(图2B)。因此,天门冬苷信息素对配偶偏好的性别歧视方面是非必需的。但它能助力雄性进行配偶选择,当雄性在daf-22突变雌雄同体线虫与野生型雌雄同体间选择时,更倾向于选择野生型雌雄同体(图2C)。
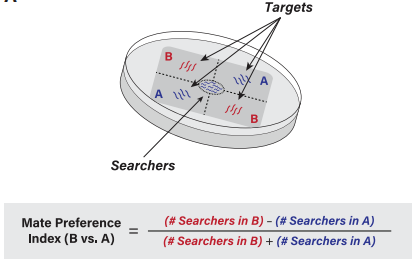
图1 配偶偏好指数统计实验方法
将两类均携带unc-54(e190)的线虫,分别置于A、B象限中,30min后将10只雄性置于平板中心,在接下来的90min内,每间隔30min记录雄性所在位置,计算每个时间点的配偶偏好指数值,平均这些数据,以获得最终的配偶偏好指数

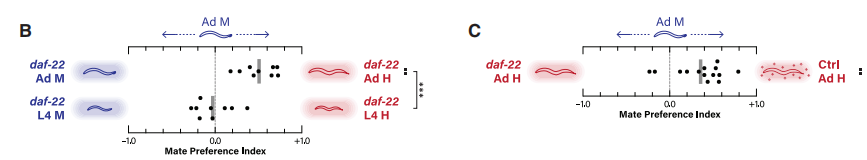
图2 daf-22突变对雄虫的配偶偏好的影响
当然,天门冬苷产生的性别特异性取决于肠道的两性状态。天门冬苷是在肠道中产生的,其中过氧化物酶体的差异表达被认为是导致天门冬苷产生性别二型性的原因。那么,肠道的性别状态会影响潜在配偶间的吸引力吗?为深入探究此问题,作者构建了两种线虫株,一种是intmasc雌雄同体,其肠道的性别状态通过雄性性别调节因子fem-3的外源性表达而被诱导呈现雄性化特征。 另一种是intfem雄虫,其肠道中表达了雌雄同体性别调节因子tra-2而被诱导呈现雌性化(图3F)。通过配偶偏好分析可知,雄性线虫更喜欢野生型雌雄同体,而非intmasc雌雄同体。并且这种偏好性具有daf-22依赖性(图3H)。而雄性对野生型雄虫与intfem雄虫没有明显偏好性,这可能是雄虫肠道的雌性化不足以触发雌雄同体典型的信息素导致的(图3J)。
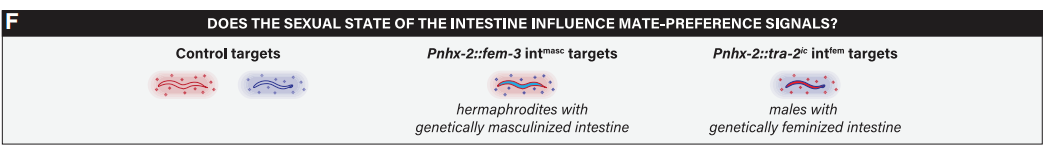
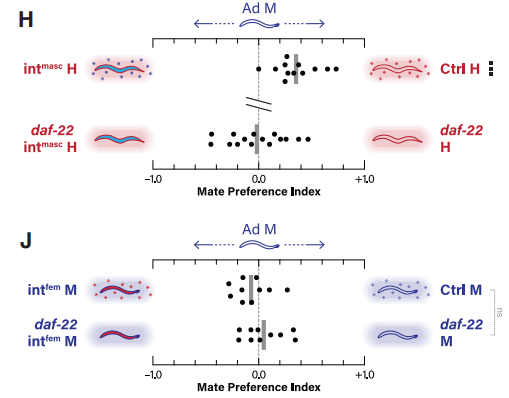
图3 雄虫对肠道异性化线虫的偏好差异
影响因素二:雌雄同体的角质层
在探讨雄性识别雌雄同体机制的复杂性时,接触相关的化学与机械线索被视为重要的调控因素。之前研究发现,雄虫与角质层蛋白的糖基化缺陷的雌雄同体的接触时间会更少,这提示表面糖蛋白可能在配偶偏好中起作用。
bus-2是一种糖基转移酶,与表面蛋白的o-联糖基化有关。雄虫对携带bus-2突变的雌雄同体线虫的喜好明显低于野生型同体线虫(图4D-E)。当在皮下特异性表达bus-2,可以挽救bus-2突变的吸引力表型(图4A)。并且,daf-22; bus-2双突变体的吸引力明显低于单个突变体(图4G),这不仅表明两基因在促进吸引力方面各自独立运作,而且与它们各自独特的生化功能及作用位点相吻合。由此可知,天门冬苷和bus-2依赖的表面线索协同优化雄性配偶偏好。交配前信号,如接触反应,作为雄性交配行为的第一步,似乎特别容易受到表面线索变化的影响。作者发现,与对照组的雌雄同体相比,雄性对bus-2表现出明显较少的接触反应行为(图4H),相反的,daf-22的缺失并没有显著减少接触反应行为(图4H),这与天门冬苷信息素通过影响配偶偏好的早期步骤来影响配偶偏好行为的观点一致。
同样地,皮下组织的性别状态控制着性别特异性表面线索的产生。将雄虫的皮下组织进行雌性化后, hypfem雄虫与野生型雌雄同体一样,具有留住雄虫的能力。
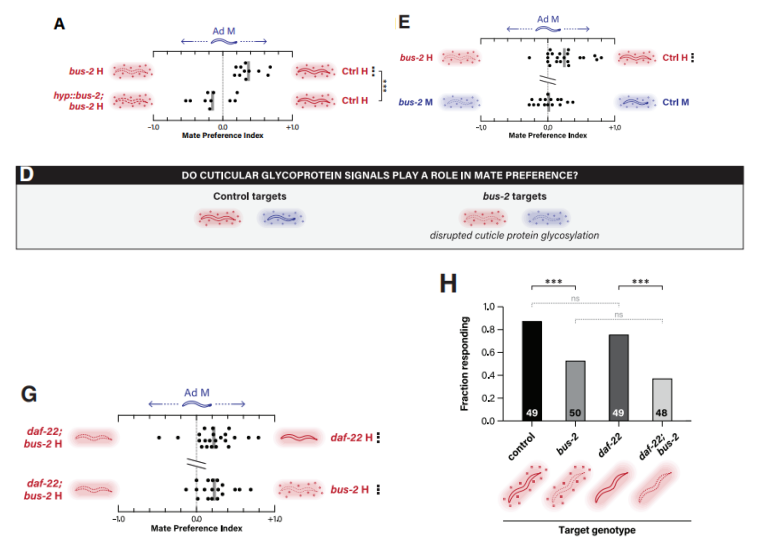
图4 表面线索对雄虫吸引力的影响
影响因素三:具有挥发性的性信息素
当雌雄同体线虫的体内精子耗尽时,便会产生挥发性的性激素(volatile sex pheromones,VSPs)。虽然VSPs的分子性质尚不清楚,但它们的产生不依赖于daf-22,而且它们不太可能与天门冬苷有关。雄性能够通过化学受体srd-1检测VSPs,该基因在性别共享的AWA嗅觉神经元中特异性表达。
基于老年unc-54雌雄同体的体内会保留卵子并变得不健康,不适合进行配偶偏好分析,故作者采用glp-1突变体,该突变体在年轻成虫时即可产生VSPs。与对照组的雌雄同体相比,雄性表现出对glp-1雌雄同体的显著偏好。雄虫对glp-1雌雄同体吸引力的增加完全依赖于其srd-1的功能(图5A-B)。即使将glp-1和对照的雌雄同体线虫置于放有雄虫的平板上方3mm处,雄虫依然明显聚集在glp-1线虫这一侧(图5C)。不过,这种效应仅限于30分钟的时间点,这表明雄性可能会迅速适应VSPs,并在没有其他线索留住它们的情况下,会离开glp-1线虫所处的位置。由此得知,VSPs能从远处吸引雄虫,而天门冬苷和/或表面线索可能对近距离留住雄虫是至关重要的。
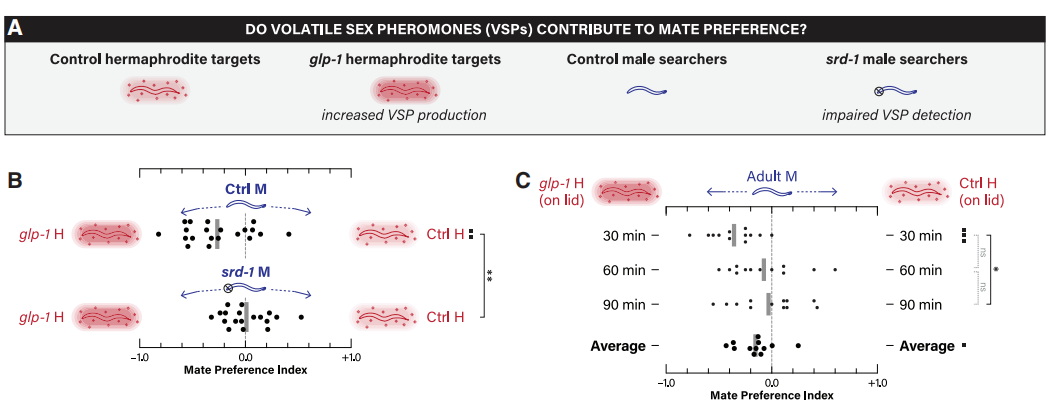
图5 VSPs在雄性对配偶偏好中的作用
影响因素四:雌雄同体的营养状况
雌雄同体的营养状况强烈地影响卵母细胞的质量和数量。在喂养良好和饥饿4小时的成年雌雄同体间,雄性没有明显的偏好。然而,雄性明显更喜欢喂养良好,而非经历28小时饥饿的雌雄同体(图6)。同样地,饥饿的L1幼虫增加了osas#9的产生(一种产生厌恶的天门冬苷),降低了ascr#3水平(一种产生吸引力的天门冬苷)。作者推断,可能是由非短链天门冬苷信息素介导了长期禁食的厌恶效应。
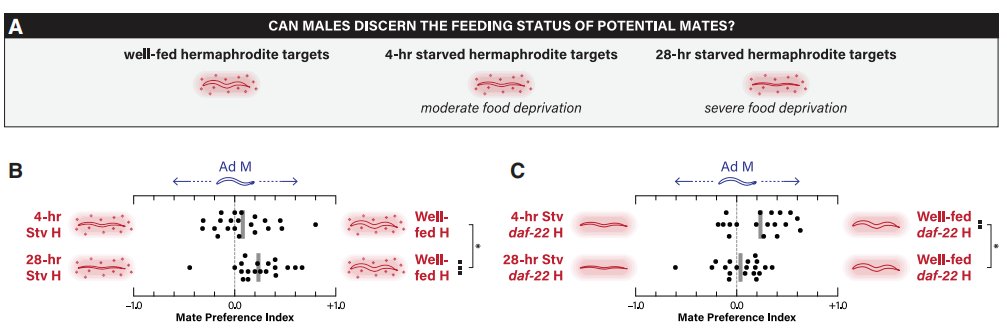
图6 雄性对不同营养状态雌雄同体的偏好性差异
影响因素五:雌雄同体的交配史
未交配和已交配的成年雌雄同体都是可育的,然而,雄性可能更喜欢未交配线虫,这样它们的精子就不会与其他雄性精子发生竞争。先前的研究表明交配降低了无精子的fog-2雌雄同体产生的VSPS效力。本研究中发现,雄性表现出对未交配雌雄同体的明显偏好(图7E),这种能力并不依赖VSPs化学受体srd-1(图7F),以及雌雄同体bus-2的功能(图7H),但是,在daf-22突变的雌雄同体中,未交配线虫的吸引力明显降低,由此说明交配的雌雄同体的天门冬苷生物合成或释放发生了变化,在没有天门冬苷信息素的情况下,雄性在检测交配状态方面的能力显著下降(图7G)。
交配史的影响表现在以下两方面:其一,对于野外分离的线虫,交配后雄性会在雌雄同体体内放置交配塞,以此降低雌雄同体的吸引力。实验室培养的线虫,虽然丧失该功能,但仍然可以在雌雄同体身上留下它们相遇的证据。其二,交配会损害雌雄同体的角质层,使雌雄同体表面产生厌恶线索。交配后的雌雄同体吸引力的下降可能取决于雌雄同体的天门冬苷和交配后的厌恶线索之间的相互作用。
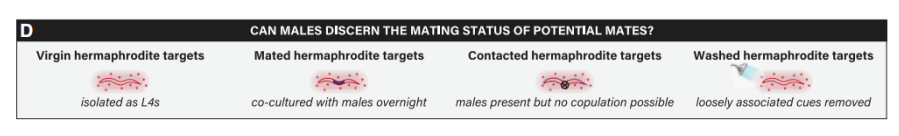
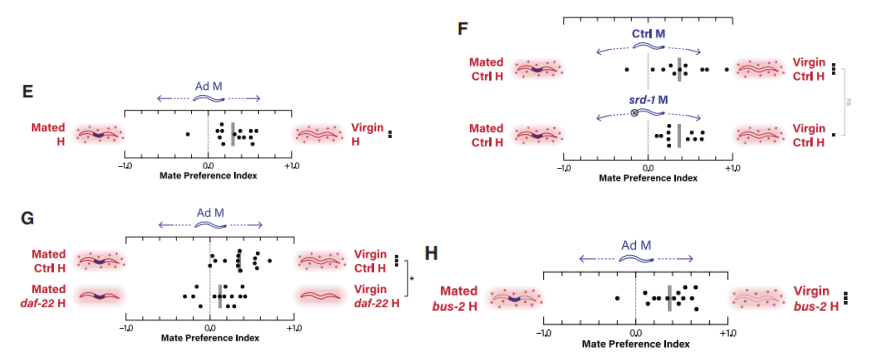
图7 daf-22依赖性的交配史对雄虫吸引力的影响
作者深入探究了配偶偏好行为的机制,发现性别共享神经元的雄性特异性特征,要么是共享的感觉神经元,要么是中间神经元接收来自雄性特异性感觉神经元的输入。由于多种感觉神经元与天门冬苷的检测有关,通过雄虫单神经元细胞消融实验得知,ADF、ASH分别消融后,均严重损害雄虫对野生型雌雄同体(与daf-22突变相比较)的偏好性(图8)。提示ASH生理学和/或连通性的重塑可能使它能够介导雄性的性欲反应。
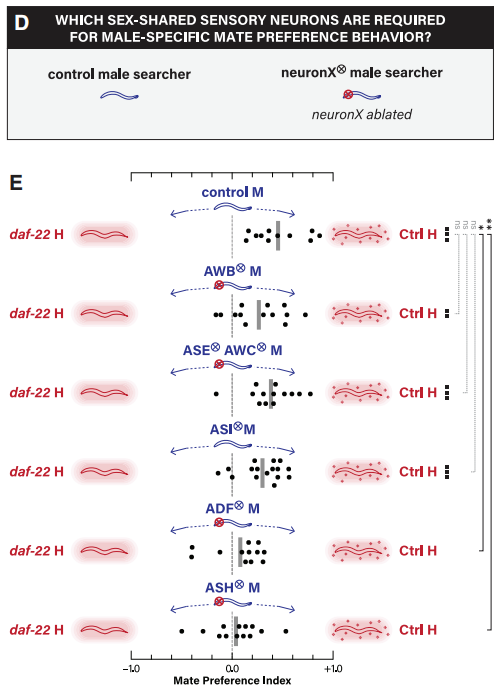
图8 单神经元细胞消融对配偶吸引力的作用
在脊椎动物中,使用多模态线索对配偶选择决策是很普遍的现象,并且这些线索的使用遵循着明确的等级序列。虽然在本研究背景下,作者并不认为线虫雄性在使用配偶偏好线索时存在一定的等级制度,但无可争议的是,动物行为学中普遍存在着利用多种信号综合判断配偶质量的策略(图9)。线虫存在一种“自交综合征”(有性生殖特征的退化),然而,种群遗传学研究揭示了其野生种群中零星杂交的证据。因此,配偶偏好行为可能是杂交受精对种群适应度贡献的一个重要决定因素,特别是在应激时期。作者的研究发现,雄性将雌雄同体的进食和交配状态准确地整合到配偶选择决定中,至少部分被雄性所采用的线索直接关联于潜在配偶的健康状况。雄性如何将这些指标融入他们的行为中,以及雄性自身的生理状态和生殖经验如何影响配偶的选择,这些都是这些研究为未来的研究提出的许多有价值的主题之一。
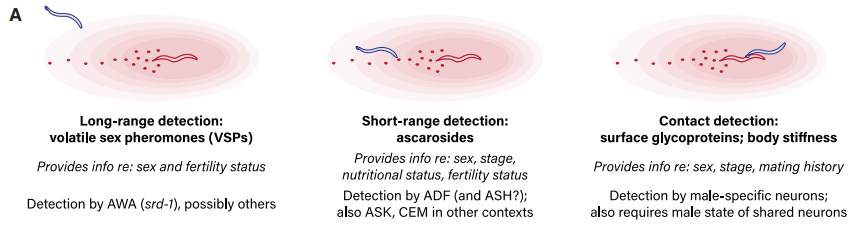
图9 不同的配偶偏好信号的物理属性示意图
撰稿:郑慧
审核:陈岚彬
编辑:余雯

