
点击蓝字
,关注我们
生物体通过内部代谢信号与外部信息的协同作用,精确感知环境食物状况,对动物生存至关重要。在秀丽隐杆线虫中,环境刺激(尤其是食物)通过嗅觉与味觉系统被感知,引导线虫向食物源移动。接触食物后,线虫减缓移动并延长停留时间;食物耗尽时,则加速并启动局部搜索行为(local search behavior,如Ω形转弯/omega turn、反向运动),随搜索时间延长转为全局搜索(global search),以更广泛范围觅食。这些行为受时间与代谢状态调控,引导线虫维持能量稳态。
食物剥夺引发能量不足,影响动物行为决策。本质上说,能量稳态部分依赖于AMPK(AMP-activated protein kinase)的调控,与其他不同的代谢信号通路协同作用。AMPK激活促进食物搜索,其废除则消除此响应,但上游信号机制尚不明晰。食物代谢涉及复杂酶促网络,最终通过线粒体氧化磷酸化(oxidative phosphorylation,OxPhos)生成ATP。此过程中,电子传递链(electron transport chain,ETC)复合物I、III和IV驱动质子泵出,形成质子电动势差(protonmotive force,PMF),进而驱动ATP合成。在功能作用上,PMF不仅供给能量,还参与调控钙信号、氧化还原信号、线粒体形态等,AMPK则通过磷酸化线粒体分裂因子发起线粒体分裂,并加速招募原分裂动力相关蛋白1(DRP-1)。
尽管PMF对细胞功能至关重要,但其生物体感知机制及信号通路间交流仍待阐明。PMF动态变化反映线粒体对生理与病理信号的响应,其降低与多种疾病相关,如缺血性疾病、神经退行性疾病等。在线虫中,肠道食物摄入反映代谢状态,神经元感知此状态并调节行为。然而,现有工具,以线粒体质子载体(protonophores)为例,如碳酰氰-4-三氟甲氧基苯腙(carbonyl cyanide p-trifluoromethoxyphenylhydrazone,FCCP,可有效地瓦解PMF,解偶联ATP生成)缺乏组织特异性与可逆性,限制了对PMF动态调控的深入理解。同样地,遗传工具,比如过表达解偶联蛋白,可引起大规模的代谢重塑,但这可能对急性信号通路产生的实验结果的诠释能力有限。
在本次的论文深入解读中,作者为了解决上述这些限制,使用光遗传学的方法时空性控制PMF,评估其在调节跨组织信号中发挥的因果作用。罗切斯特大学Andrew Wojtovich团队发现,肠道而非神经元PMF降低驱动觅食行为变化,且神经元AMPK调节此响应。由此产生的神经元PMF损失主要通过AMPK在5-羟色胺能或多巴胺能神经元中传递,影响全局搜索行为。研究还探讨了线粒体亚细胞定位与动态融合-分裂在线虫全局搜索中的作用,揭示了以线粒体为中心的跨组织信号轴,整合能量感知与代谢信号协调。


PMF消散诱导线虫从局部到
全局搜索的行为转变
在食物剥夺一段时间后,线虫的觅食行为由局部转变为全局搜索模式,然其分子调控机制尚未明晰。全局搜索行为的表现特征包括增加线性正向运动、降低Ω转弯和反向运动(图1A)。鉴于食物剥夺可能影响能量代谢,作者首先探究了食物匮乏引发的内部代谢信号对线粒体膜电位(PMF)的影响。作者使用TMRM(四甲基罗丹明甲酯,TMRM可以快速通过细胞膜,仅需几分钟就可被具有活性的线粒体所俘获,并且对细胞没有毒性。其荧光的增强或减弱说明线粒体膜电位的变化,线粒体膜电位的高低变化决定了TMRM的分布浓度。电位高,TMRM在线粒体的浓度高,显示为强力红色或橘红色;反之,TMRM弥散在胞浆内,荧光减弱表明线粒体膜电位降低)衡量PMF,发现在食物剥夺后TMRM荧光减弱。为评估PMF在食物相关行为中的作用,作者利用药物FCCP来瓦解PMF,抑制氧化磷酸化产生的ATP。确保在暴露处理24 h后药物全身组织分布,移除食物后评估觅食行为。结果发现FCCP处理后降低了Ω转弯和反向运动的频率,重现了线虫行为从局部搜索到全局搜索的时间依赖性变化(图1B)。为排除细菌代谢干扰,使用热杀死细菌喂食亦得相同结果。这些数据表明PMF作为一种信号,对线虫的行为转变产生影响。
行为响应是一个动态的过程,使用药物处理缺乏瞬时分析信号通路和评价细胞特异性的能力。为了克服该限制,作者使用线粒体靶向的光遗传学工具mtOFF(mitochondria-OFF),mtOFF是遗传编码的光激活质子泵,它能单向地能泵出质子从而去极化PMF。与使用FCCP对线虫处理的结果相似,mtOFF激活可瓦解PMF,并导致降低ATP水平和补偿性的增强ETC活性。但是mtOFF可靶向特定的组织,能同光照极性地激活/去激活PMF消散,达到更强的时空精确性。mtOFF激活过程需要全反式视黄醛(all-trans-retinal,ATR)作为辅助因子,但又因线虫自身无法合成ATR,所以激活mtOFF的外部条件需要同时添加ATR和光照。与长时间的食物剥夺和FCCP处理线虫的结果相似,mtOFF激活导致降低线虫Ω转弯和反向运动(图1C)。
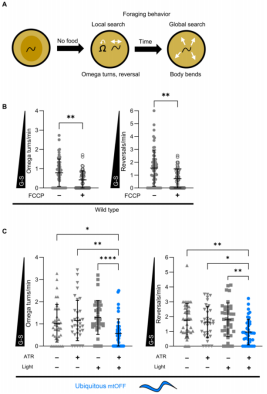
图1 PMF消散改变线虫从局部到全局搜索的觅食行为
肠道代谢状态控制觅食行为
线虫的肠道是吸收和存储营养的器官,而神经元能感应食物存在与否。相应地,作者构建在线虫肠道(vha-6启动子)和神经系统(rab-3启动子)中表达mtOFF的品系,用于组织特异性的控制PMF消散。实验首先确认组织特异性的光遗传消散PMF是否影响细胞内的能量水平。正如前文提到的,使用泛表达的mtOFF控制PMF消散增加补偿性的氧消耗率(oxygen consumption rates,OCRs)和减少全身ATP水平。在肠道和神经元中表达mtOFF时,作者检查了线虫全身的氧消耗率。试验结果显示只有当肠道中mtOFF激活时,氧消耗率才会增加(图2A),而神经元中mtOFF激活时并不能(图2B)。而生物发光法检测组织特异性PMF消散的线虫全身ATP产量的结果显示,当肠道PMF消散时会降低全身ATP水平(图2C),神经元中激活mtOFF并不能改变全身ATP水平(图2D)。上述肠道内ATP水平降低和氧消耗率增加的实验结果与PMF消散是相一致的。
在线虫中,肠道组织占据全身更大的比例。作者推测可能在神经元中mtOFF激活而降低ATP水平不足以影响到线虫全身ATP水平。为了解决这个问题,作者使用基于荧光共振能量转移(fluorescence resonance energy transfer,FRET)原理设计的ATP生物感受器——ATeam,ATeam可以原位检测细胞内的ATP水平(图2E),分别在肠道和神经元中表达ATeam,ATeam的发射光谱与mtOFF激发波长相重叠。但是从对照组实验发现ATeam信号并不能激活mtOFF,这表明活体动物中具有细胞类型特异性的独立监测和调节能量平衡的能力。更进一步的,使用相同的肠道特异性启动子(vha-6)表达ATeam和mtOFF对mtOFF的水平没有影响。借助ATeam在线虫体内的表达,作者观察到在mtOFF激活后,肠道和神经元中ATP水平降低(图2F、G)。这个实验证实了靶向光遗传PMF消散可降低组织特异性的ATP水平。
为了更进一步测试组织特异性PMF改变如何被感知并体现在食物相关的行为,作者定量测量了线虫在急性食物剥夺后的觅食行为。结果发现,通过组织特异性的mtOFF表达来限制PMF在肠道的耗散,导致了线虫向全局搜索行为的转变(图2H)。尽管神经功能的主要作用是控制行为,但是神经元中PMF消散并不能引起有效的全局搜索行为改变(图2I)。虽然肠道中PMF消散驱动了觅食行为,但是行为本身是一个涉及不同组织信号相互协调的过程。
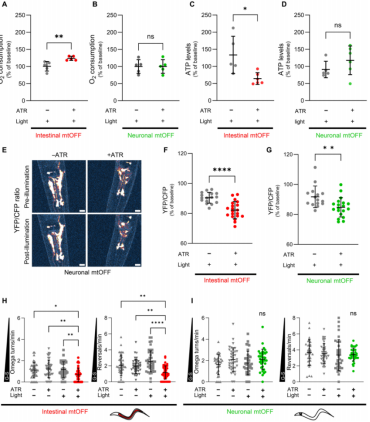
图2 组织特异性的PMF消散降低体内的ATP水平
神经元的AMPK对PMF诱导的
全局搜索行为是必需的
降低肠道PMF能有效地转变线虫食物搜索行为。为了进一步探究识别传递PMF消散信号的分子组分,作者测试了AMPK是否在PMF诱导的行为变化中发挥关键作用。AMPK被认为是细胞能量的传感器,将生物能量生成和生理响应与细胞能量状态相互耦合。多种细胞输入信号,例如AMP/ATP比率、ROS和PMF消散,都能引起AMPK激活。作者使用泛表达mtOFF和AMPK的α-催化亚基aak-2(与人源PRKAA1和PRKAA2同源)突变体aak-2(ok524)来研究AMPK与PMF诱导的行为之间的关系。结果表明,aak-2突变体可有效地压制因全身PMF消散产生的全局搜索行为表型(图3A)。尽管肠道中PMF消散能有效地发起行为响应,但是再表达AMPK却不能挽救这种表型(图3B)。这些都表明,存在肠道以外的跨组织信号轴调节线虫行为。
已知线虫神经系统能感知食物,作者使用两种不同的泛神经元表达启动子(unc-119和Ultra Pan-Neuronal,UPN)在aak-2突变体中再表达AMPK。试验结构显示,两种启动子均能挽救PMF引起的反向运动频率降低,但是并不影响Ω转弯频率(图3C、D)。这暗示Ω转弯并不是受AMPK调节的能量感知行为。这些发现总体上表明肠道PMF信号流以跨组织方式通过神经元中的AMPK促进觅食行为。
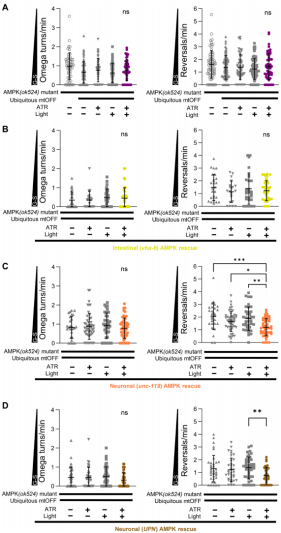
图3 神经元中AMPK对于觅食决定的PMF信号是必需的
尽管存在食物,在5-羟色胺能和多巴胺能
神经元中PMF-AMPK信号耦合引起行为改变
在食物缺失条件下,PMF是线虫从局部到全局搜索模式转变中重要的信号。作者测试发现,即使在存在食物条件下,PMF消散也能影响线虫行为。上文中我们讲到,线虫在食物中运动更慢并且停留的时间更长,运动降低是通过计算在食物中的身体弯曲(body bends)次数评定的。作者发现暴露于FCCP的线虫在食物存在下也增加身体弯曲频率,这个结果与食物剥夺产生的行为效果是一致的,而这个行为我们定义为全局搜索(图4A、B)。为了排除细菌对FCCP代谢的影响,作者在实验中再次使用热杀死的细菌喂食线虫,仍然观察到增加身体弯曲频率的现象。而且,相关研究工作同样表明,AMPK缺失废除PMF诱导的全局搜索表型,且通过泛神经元中表达AMPK能挽救该表型。这些结果表明,即使在食物可获取情况下,与PMF耦合的AMPK在自发调节觅食行为中的组织特异性作用。
上文中作者揭示了神经元AMPK对PMF调节的行为响应是必需的,线虫具有多种类型的神经元,接下来作者想要测试哪一类型的神经元负责驱动该效果。本研究在aak-2突变体背景下,使用细胞类型特异性的启动子再表达aak-2,从而筛选出不同神经元类型在AMPK调节的食物行为中的作用。首先确认在aak-2突变体背景中使用泛神经元启动子表达AMPK可增加线虫的运动(图4C)。在涉及线粒体功能【如ROS信号,ASH神经元(sra-6启动子)和I2中间神经元(flp-15启动子)】以及与食物相关行为【5-羟色胺能神经元(tph-1启动子),多巴胺能神经元(dat-1启动子),AIY中间神经元(ttx-3启动子),AIB中间神经元(npr-9启动子),AVA中间神经元(rig-3启动子)和AIA中间神经元(gcy-28.d启动子)】的中间神经元中再表达AMPK,结果显示在AIY,I2,AIB,ASH,AVA,AIA中再表达AMPK不能有效地挽救全局搜索表型,但是在5-羟色胺能神经元和多巴胺能神经元中表达AMPK能挽救线虫的运动速度(图4C)。AMPK与5-羟色胺能和多巴胺能神经元回路之间存在联系,特定神经递质的神经元类型与线粒体能量感知之间存在偶联以控制线虫行为。
有研究表明神经递质(5-羟色胺和多巴胺)具有调节线虫食物相关的行为。上文中作者验证了在5-羟色胺能和多巴胺能神经元中再表达AMPK,可在存在食物条件下增加线虫身体弯曲频率(图4C)。接下来,作者测试在单个细胞类型中PMF消散是否能如之前使用泛神经元再表达AMPK一样,有效地引起食物剥夺的觅食表型。AMPK在5-羟色胺能和多巴胺能神经元中的挽救都不能恢复PMF诱导的反向运动频率的降低(图4C、D)。为了比较不同基因型线虫品系对mtOFF激活产生的行为响应,作者首先对反向运动和Ω转弯频率做归一化处理,即控制条件mtOFF激活(+ATR +light)与mtOFF未激活(−ATR +light)做比较。这种数据处理有助于排除由于基因型变化造成的基础行为的差异性,并允许直接比较突变体或挽救基因型线虫品系之间相对于mtOFF激活之间的效应。当AMPK在5-羟色胺能或多巴胺能神经元中再表达时,mtOFF对觅食行为的影响是相似的(图4F、G)。总之,比较相对变化反映出在肠道中mtOFF激活和挽救AMPK功能很大程度上恢复了局部搜索响应(图4F、G)。文章的数据表明PMF诱导的身体弯曲和局部搜索行为参数(Ω转弯和反向运动)改变是与不同的神经元类型之间产生相互交流的。即使在食物存在条件下,由PMF消散引起的神经元AMPK激活能也可以有效地改变线虫行为,但在能量缺乏的环境中则不会。
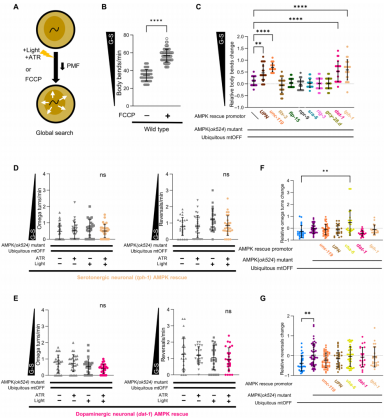
图4 5-羟色胺能和多巴胺能神经元中的AMPK上调PMF消散调节全局搜索
线粒体动态平衡对于食物相关的行为是必需的
线粒体是动态变化的,它会随着代谢和细胞应激变改变形态。此外,AMPK的磷酸化作用触发线粒体分裂机制。因此,作者想要测试线粒体分裂是否会加速食物相关的行为。在drp-1(tm1108)缺失突变体背景中,作者观察到线粒体分裂功能缺失抑制PMF依赖的增加身体弯曲的表型(图5A,与图4B比较)。相似地,PMF消散没有影响反向运动和Ω转弯的频率(图5A、B)。文章的数据表明线粒体分裂在PMF感知食物相关行为中发挥重要作用,因为通过PMF消散引起的食物搜索行为被线粒体分裂功能缺失抑制了。接下来,作者使用AMPK激活剂(5-aminoimidazole-4-carboxamide ribonucleotide,AICAR,是一种AMP类似物)检查AMPK与线粒体分裂之间的关系。文章发现使用AICAR处理drp-1突变体不能影响在食物中的线虫身体弯曲或反向运动和Ω转弯的频率,该结果暗示AMPK激活信号位于线粒体分裂的上游,这与AMPK诱导线粒体分裂的作用是一致的。
在急性食物剥夺条件下,线粒体倾向于融合,这可能与增强氧化磷酸化活性有关。作者推测融合可能涉及PMF消散的细胞感应。为了验证上述推测,文章将fzo-1(tm1133)(人源MFN2同源蛋白)与泛表达mtOFF转基因线虫杂交。在缺失线粒体融合功能下,mtOFF激活不能改变弯曲次数(图5C)或Ω转弯和反向运动频率(图5D)。因此,在无线粒体动态变化(分裂-融合)情况下,PMF消散导致的身体弯曲增加表型被忽略了(图5E)。尽管由PMF消散诱导的觅食行为受线粒体融合缺失的影响要大于线粒体裂变的影响(图5F)。在融合突变体中Ω转弯和反向运动的基线频率要更低,这说明无论是否PMF消散,线粒体融合缺失都废除了线虫对食物的感知能力。这些都说明,线粒体动态失衡压制PMF消散对感知食物获得的能力。
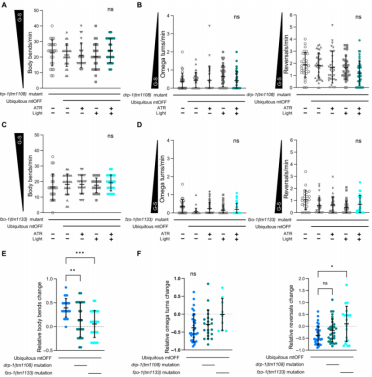
图5 线粒体动态变化调节线虫的觅食行为
神经元中完整的线粒体转运对于感知
PMF诱导的全局搜索行为变化是必要的
除了上文中提到线粒体分裂-融合的动态外,将线粒体转运到神经元末梢也是线粒体动力学的重要内容,正确的线粒体定位对于发挥神经元的功能和信号传导是至关重要的。线粒体的亚细胞定位影响神经元中线粒体的特殊形态,并标明其功能改变。因此,作者测试神经元中线粒体定位对于PMF诱导的行为改变是否是必需的。文章中使用ric-7(n2657)突变体,该突变体缺失神经元线粒体从细胞体向轴突运输所必需的衔接蛋白(图6A)。因此,在轴突无线粒体的背景且PMF消散时,线虫在食物中的移动速度并没有改变。接下来,文章使用嵌合蛋白Mitotruck【Mitotruck是由驱动蛋白-1(线虫中的是UNC-116)与线粒体外模蛋白Tom7(线虫中的是TOMM-7)组成的嵌合蛋白】挽救线粒体运输缺陷的表型。结果发现Mitotruck可以挽救PMF诱导的全局搜索表型(图6C、D),证实了位于轴突的能量供应对于行为响应的重要性。
文章向我们展示食物存在条件下,神经元轴突中的线粒体在传递线虫行为信号中的重要性。那么,神经元中线粒体定位对在食物缺失条件下是否是必需的呢?与身体弯曲的试验结果一致,作者并没有观察在线虫轴突缺乏线粒体的背景下由PMF消散引起的局部搜索行为差异(图6E)。但是线虫轴突缺乏线粒体降低了线虫基线觅食行为,这表明在此背景下干扰了线虫对食物感知的能力。更进一步地讲,表达Mitotruck不能有效地恢复由PMF诱导的反向运动和Ω转弯频率(图6F)。相应地,在线粒体转运受损和挽救线虫中,由PMF诱导的Ω转弯的变化程度相对较小(图6G)。反向运动频率在PMF消散中并没有发生改变(图6H)。总体来讲,该数据强调了线粒体PMF在能量状态与影响行为沟通方面扮演着重要的媒介作用。
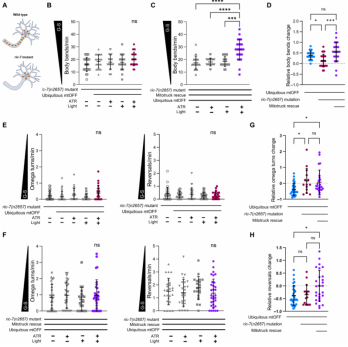
图6 在饲养良好的线虫中,轴突线粒体完整的转运动能对PMF诱导的速度升高是必需的
小编总结
线虫为了食物探索它们所在的周围环境,食物的可获得性影响线虫的觅食行为,这些行为又影响着线虫发现并停留在食物源的可能性。作者使用光遗传学的方法消散PMF并识别这个过程产生的信号通路。使用遗传和药理技术相结合的技术,文章向我们展示组织特异性的代谢状态是如何改变线虫的觅食行为。在食物缺失情况下,线虫的行为模式由局部搜索向全局搜索转换,增加线虫搜索的范围,这种行为模式的改变是受到肠道PMF选择性驱动的。而且,神经元中AMPK和轴突线粒体转移功能涉及对内部代谢状态的感知。
文章中大量应用到的mtOFF瓦解线粒体PMF,该光遗传操作方法是Andrew Wojtovich研究团队在2020开发的(Berry BJ, et al. FASEB J. 2020;34(12):16333-16347.),同年他们又开发出mtON(mitochondria-ON,Berry BJ, et al. EMBO Rep. 2020;21(4):e49113.),相对于本文中介绍的mtOFF对PMF消散,mtON通过光遗传方式可逆地对PMF强化。联合mtOFF和mtON方法,我们可以实现对细胞中的能量工厂——线粒体PMF进行组织特异性的精确操作。
Reference
Vodičková A, Müller-Eigner A, Okoye CN, et al. Mitochondrial energy state controls AMPK-mediated foraging behavior in C. elegans. Sci Adv. 2024;10(16):eadm8815. doi:10.1126/sciadv.adm8815
撰稿:卞文印
审核:陈岚彬
编辑:余雯

