

点击蓝字 关注我们

在衰老过程中,环境和遗传因素均起到了重要的作用。虽然极端寒冷的环境对生物体有害,但适度的体温降低确实能显著延长寿命。因此,降低体温是延长生物体寿命的有效措施之一。除了温度因素外,繁殖力也与寿命呈负相关。大量研究表明,在标准(20℃)和温暖的温度下,线虫的性腺会产生某种信号,促进体细胞的进行性恶化和机体老化。缺乏性腺的线虫寿命可延长60%,并对包括高温在内的各种环境压力表现出更强的抵抗力。然而,有研究发现低温并不能延长缺乏性腺线虫的寿命。那么,性腺在衰老中究竟扮演了怎样的角色呢?让我们从“Prostaglandin signals from adult germline stem cells delay somatic aging of Caenorhabditis elegans”文中一起寻找答案吧。
明确性腺对低温诱导长寿表型的重要性
线虫性腺由远端有丝分裂区、从减数分裂早期到成熟配子的细胞区域组成,并向近端延伸。成虫生殖细胞在配体发生和细胞死亡过程中不断丢失,而有丝分裂区增殖的生殖干细胞(GSCs)会自我更新,并提供进入减数分裂的新细胞,进而产生配子(图1)。
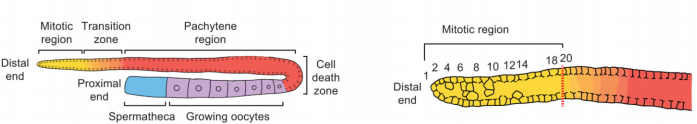
图1 年轻成虫雌雄同体性腺示意图
德国科隆大学David Vilchez团队首先检测了glp-1(e2141)突变线虫,这些线虫由于在限制性温度(25℃)发育过程中生殖细胞增殖失败而发育成缺乏生殖性腺的成虫。研究人员在25℃培养glp-1(e2141)突变体的幼虫以获得缺乏性腺的成虫,随后将其转移到不同的温度下进行后续观察。与相同条件下的野生型动物相比,缺乏glp-1性腺的线虫在20℃下更长寿。相反,在较低温度下(即15℃,10℃)下,glp-1突变体的寿命明显短于野生型线虫。
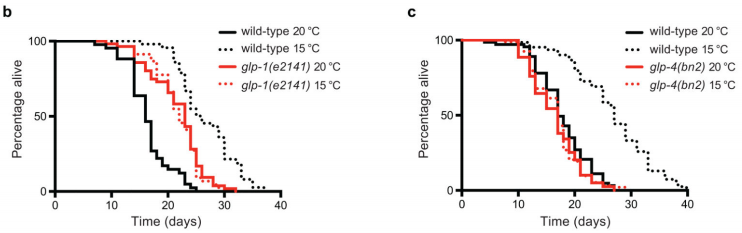
图2 性腺缺乏突变体在不同温度下的寿命情况
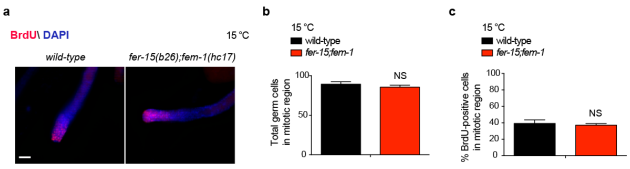
图3 fer-15;fem-1不孕突变体的有丝分裂区生殖细胞数量比较
低温延缓了GSCs的衰竭和生殖老化
同样地,作者在缺乏性腺的glp-4(bn2)突变体线虫中发现,无论是在20℃还是15℃条件下,实验结果均显示未能延长该线虫寿命。然而,低温下处理fer-15(b26);fem-1(hc17)不孕线虫株(在发育过程中,在限制性温度25°C下培养时,该线虫株性腺可产生生殖细胞,培养至成虫后转移至低温)却能增加其寿命。因此,研究人员得出结论:寒冷诱导的长寿表型的缺失并非由于不育引起,而是由于性腺增殖的耗竭。性腺在成年期对寒冷诱导的长寿表型尤为重要(图2-3)。
为了确定温度和年龄对成虫性腺增殖的影响,作者在20℃培养线虫直到成年,随后转移至不同温度下。值得注意的是,有丝分裂区域内生殖细胞总数并不会随年龄或温度而发生变化(图4)。然而,通过溴脱氧尿苷(BrdU)标记发现,低温延缓了与年龄相关的有丝分裂细胞百分比的下降。在25℃时,生殖细胞增殖的百分比在第5天明显降低(即brdu阳性细胞小于10%),而线虫的平均寿命为14天。在20℃时,线虫的平均寿命为19.5天,在第10天生殖细胞增殖明显下降。然而,15℃条件下的成虫中,生殖细胞增殖的急性下降被延迟到第23天,更接近线虫28天的平均寿命。同样,10℃下的线虫在第37天之前都保持了很高的生殖细胞增殖比例,这批线虫平均寿命为43天(图5)。由此可知,低温环境增强了GSCs在衰老过程中自我更新的能力,有效维持了有丝分裂区域内的增殖细胞数量。
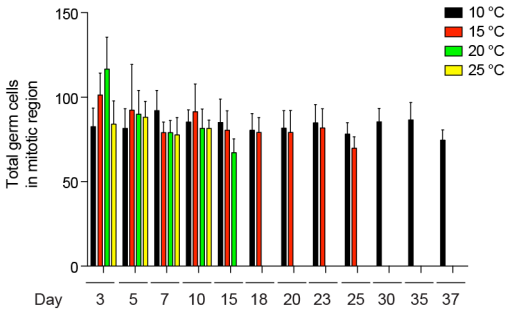
图4 野生型线虫在20℃培养至成虫后转移至不同温度下生殖细胞数量变化
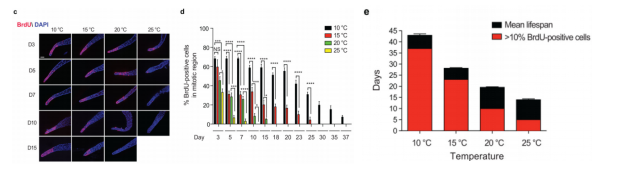
图5 野生型线虫在不同培养温度下生殖细胞的下降趋势
为了评估GSCs的维持是否与诱导冷相关长寿的能力存在关联,研究团队尝试在不同的成虫阶段将线虫从较高的温度转移到较低的温度。当线虫在20℃下培养至成虫第10天,然后转移到较低的温度时,寒冷诱导的寿命延长表型被抑制,这与20℃时性腺增殖能力的急剧下降(即<10% BrdU阳性细胞)相关。同样地,线虫在25℃下养至成年的第5天再转移至低温环境时,其低温下存活更长的能力下降,这与25℃时细胞增殖下降有关(图6)。
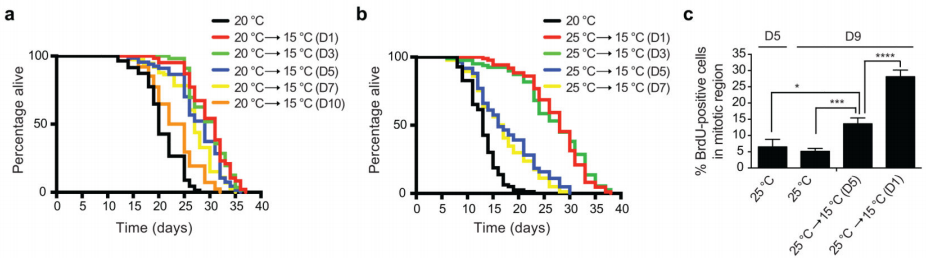
图6 不同的成虫时期从高温转移至低温后的寿命情况
生殖细胞通过诱导肠道中的
cbs-1来延长寿命
成年GSCs具备直接调节寒冷诱导寿命的能力,为研究其作用机制,研究者利用定量蛋白质组学方法及RNAi筛选,发现在生殖细胞增殖介导的冷诱导长寿过程中,需要cct亚基、hsp-1、ama-1和cbs-1的高表达。然而,关于这些因子如何调节寿命的问题仍需进一步探讨:它们是直接影响生殖细胞的增殖来调节寿命,还是通过增殖的生殖细胞在体细胞组织中诱导特定的因素,进而延长寿命?
通过后续一系列实验,结果发现ama-1缺失导致性腺增殖能力的大幅降低,仅在肠道中敲除ama-1并不影响寿命,而cct亚基和hsp-1可以同时在体细胞组织及生殖细胞增殖中发挥作用来调节机体寿命。值得注意的是,cbs-1是一个独特的寿命调节因子,它不影响生殖细胞的增殖数量。虽然在神经元中特异性敲除cbs-1并没有改变寿命,但在肌肉或肠道中下调其表达却显著缩短了寒冷诱导的寿命。
随后,作者对cbs-1进行荧光蛋白标记,以验证上述结论。实验数据显示,在20℃和25℃时,cbs-1主要在后肠中表达(图7)。在低温下,cbs-1的表达量增加,尤其是在肠道和身体肌肉中。据此得出以下结论:增殖的生殖细胞可以诱导特定体细胞组织中的cbs-1在低温下延长寿命。继而,作者在肠道过表达cbs-1,观察是否可以挽救低温下缺乏性腺线虫的短寿命。虽然在肠道中cbs-1的组织特异性过表达并没有进一步延长野生型线虫的冷诱导寿命,但它延长了glp-1性腺缺失突变体的寿命。
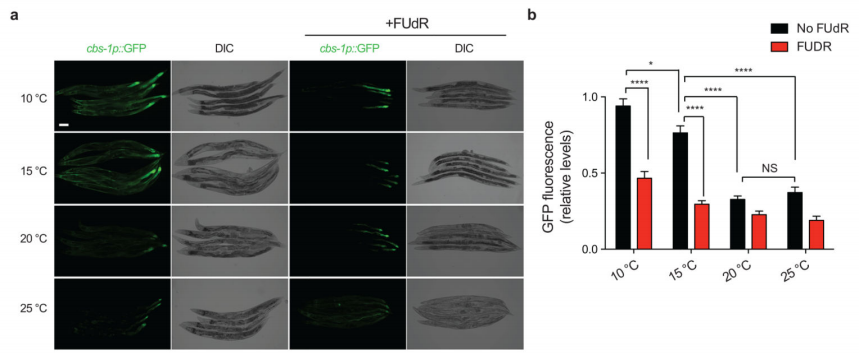
图7 GSCs在低温条件下诱导体细胞组织中cbs-1的表达
在介导半胱氨酸和同型半胱氨酸相互转化的转硫途径中,CBS发挥着关键作用。CBS活性产生胱硫氨酸和硫化氢(H2S),它作为一种气体信号分子,可以降低血压,防止哺乳动物的神经退行性变。此外,饮食限制(DR)可诱导高水平的H2S,这对酵母、线虫、果蝇和啮齿类动物的寿命延长表型极其有利。重要的是,外源添加的H2S足以延长线虫的寿命。
综上所述,由GSC增殖调节的肠道中cbs-1的上调有助于冷诱导的长寿命表型。
GSCs通过释放PGE2(前列腺素E2)
延长低温条件下的寿命
生殖细胞在低温下是如何诱导肠道中cbs-1表达的呢?作者通过在不同培养温度下进行iff-1 RNAi(iff-1在性腺中特异性表达,特别是在有丝分裂发生的远端性腺中大量表达。实验显示在成年期启动的iff-1 RNAi足以减少性腺远端生殖细胞的增殖),提取线虫性腺进行转录组分析,结果显示有248个基因上调,有433个基因下调,其中,pges-2的表达在升高温度或iff-1 RNAi中呈现一致性的下调。Western blot和免疫印迹实验表明,在低温下,PGES-2有更高的含量,且性腺较肠道有更高的表达水平(图8)。低温促使PGES-2在生殖细胞的胞浆富集,高温下PGES-2主要聚集在核内。基于该结论,他们在低温下,比较pges-2功能缺失突变株和野生型线虫株寿命,发现pges-2突变株寿命明显更短。而在20°C或25°C下,不存在明显的寿命缩短情况。同时,作者也发现:低温下pges-2突变株的体细胞中cbs-1的表达量显著下降,通过外源性补充PGE2激素,能够挽救pges-2突变株的cbs-1低表达情况及延长其寿命(图9)。
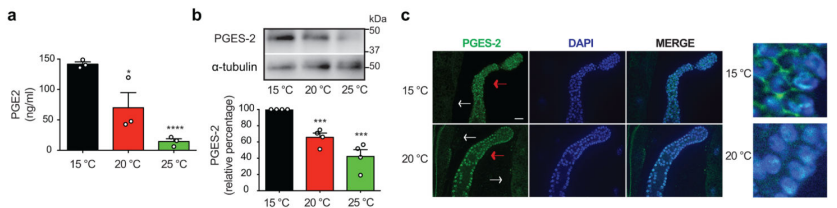
图8 不同温度下PGE2的表达差异(图8c 白色箭头指示肠道,红色箭头指示性腺)
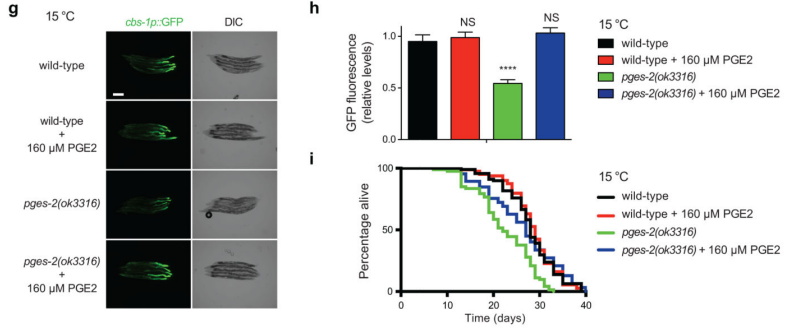
图9 PGE2对cbs-1表达量及寿命的影响
为验证由性腺释放的PGE2可以调控低温下线虫寿命,研究团队利用FUdR处理线虫(FUdR会抑制成虫GSCs的增殖),检测了添加外源性PGE2激素作用后肠道cbs-1的表达。结果显示,外源性PGE2足以挽救FUdR处理诱导的肠道cbs-1的低表达,并缓解低温下GSC增殖损失导致的寿命下降,还能挽救低温下缺乏glp-1性腺线虫的短寿命表型。综合上述数据可知,性腺通过PGE2信号与体细胞组织产生联系,以在低温下延长线虫寿命(图10)。
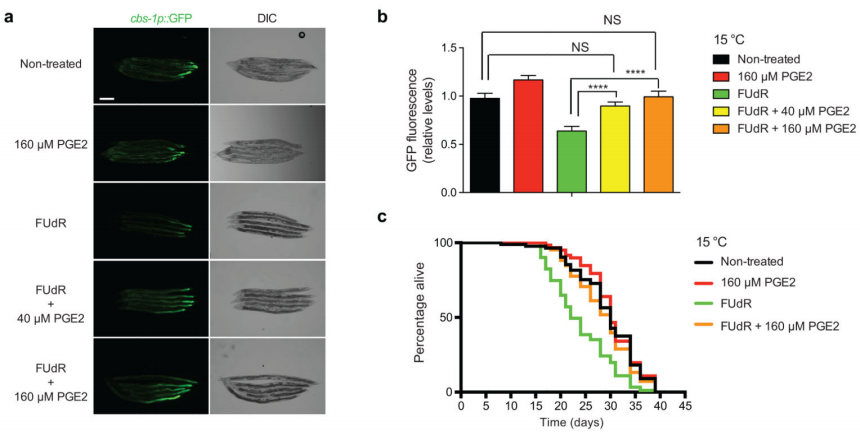
图10 性腺释放的PGE2对FUdR处理的线虫的寿命影响
总结
文章作者揭示了一种细胞非自主机制(即神经生殖系-肠轴),在低温条件下协调延长生殖能力和长寿命,而不需要牺牲性腺或体细胞(图11)。通过生殖细胞激活不同的信号来调节体细胞组织,从而在生理和环境条件发生变化时产生相应的反应。线虫GSCs这种意想不到的促长寿作用对理解衰老过程具有重要意义。
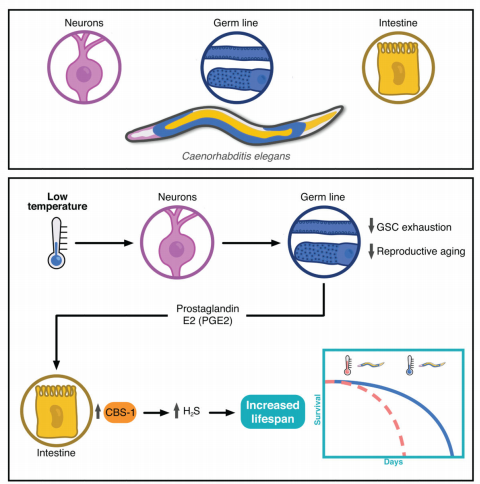
图11 低温下体细胞组织与性腺之间的细胞非自主通信模型
撰稿:郑惠
审核:陈岚彬
编辑:余雯

