

纤毛是一种基于微管结构的细胞器,它从大多数真核细胞表面突出,参与细胞运动和感觉功能。纤毛异常与人类多种遗传病相关,统称为纤毛类疾病。其组装和维持依赖于鞭毛内运输(intraflagellar transport,IFT)机制。秀丽线虫拥有的感觉纤毛是位于头部的头感器通道和尾部的尾感器通道。在线虫中通过异源三聚体驱动蛋白-II与同源二聚体驱动蛋白-2 OSM-3之间的协同作用促进向前鞭毛内运输过程,而鞭毛内运输动力蛋白CHE-3则拥有向后鞭毛内运输的能力。
最新研究揭示了纤毛在不同细胞、组织和发育阶段的多样结构和功能,特别是初级纤毛在信号路径中的关键作用。先进的成像技术促进了纤毛在神经元和昼夜节律调节中的研究。线虫感觉纤毛突变体显示长寿表型,可能与信号感知能力相关。长寿因子DAF-16/FOXO和AMPK信号通路与纤毛突变体长寿及感觉感知退化有关,但机制未明。
蛋白质稳态通过维持细胞平衡抵抗环境应激和延长寿命,内质网(ER)在其中起关键作用。打乱这种平衡会触发已知的内质网非折叠蛋白反应(Unfolded Protein Response in the ER,UPRER),其主要功能是尽可能恢复蛋白稳态和细胞平衡。依靠内质网腔传感元件不同进行划分,多细胞生物体的UPRER包含三个不同的分支:IRE1(inositol requiring enzyme 1)、PERK(double-stranded RNA-activated protein kinase [PKR]-like ER kinase)和ATF6(activating transcription factor 6)。神经系统与外周组织之间的细胞非自主的UPRER通信表明,UPRER可能参与寿命延长。
线虫tax-4和tax-2基因负责编码对感觉神经元功能至关重要的环核苷酸门控(cyclic nucleotide-gated,CNG)通道蛋白,其信号转导可能涉及G蛋白对cGMP门控离子通道活动的调节,并最终影响通道的开启或关闭。
清华大学欧光朔团队在PNAS上发表关于感觉神经元纤毛缺陷可激活非神经元的远端细胞中依赖IRE-1-XBP-1分支路径的UPRER。这种非细胞自主的UPRER现象需要神经信号,并涉及环核苷酸门控离子通道。这暗示靶向激活环核苷酸门控离子通道可能具有调节长寿的效果。现在就和小编一起学习文章的研究发现吧。

1
纤毛突变体的RNA测序(RNA-seq)反映上调表达UPRER报告基因hsp-4

为探究纤毛如何调控线虫寿命,作者对长寿表型的突变体进行转录组测序分析。研究聚焦五个纤毛突变体线虫品系,即osm-3(p802),che-3(e1124),che-11(e1810),daf-10(e1387)和osm-5(p813)(图1A)。纤毛完整性评估采用荧光染料摄入法,该方法使用的DiI染料在头/尾感器神经元积累,如线虫功能和结构完整性受损,那么线虫就无法将DiI染料摄入。所有野生型线虫可以成功地从培养基中摄入DiI染料,反之没有一株纤毛突变体可以摄入DiI染料。作者又使用GFP标签融合的IFT52/OSM-6蛋白标记纤毛,结果显示长寿的纤毛突变体其纤毛的长度显著降低了。
mRNA测序分析表明,这些突变体与野生型线虫在L1幼虫阶段之间存在明显的基因表达差异(图1B)。这五个突变体线虫转录组中有167个基因普遍上调表达(图1C、D),这些基因与纤毛的形成和维持有关(图1E),这与在纤毛受应激或突变时纤毛相关基因表达上调的结果是相一致。有趣的是,作者同时观察到UPRER基因表达上调(图1E)。线虫HSP-4是哺乳动物ER定位的Hsp70分子伴侣BiP的同源物,其在ER应激中会上调表达。IGV软件可视化显示在纤毛突变体中hsp-4表达显著上调。通过RT-qPCR分析同步化的L4阶段线虫中证实hsp-4表达升高(图1F)。因此,文章认为hsp-4在纤毛突变体确实是显著上调的。
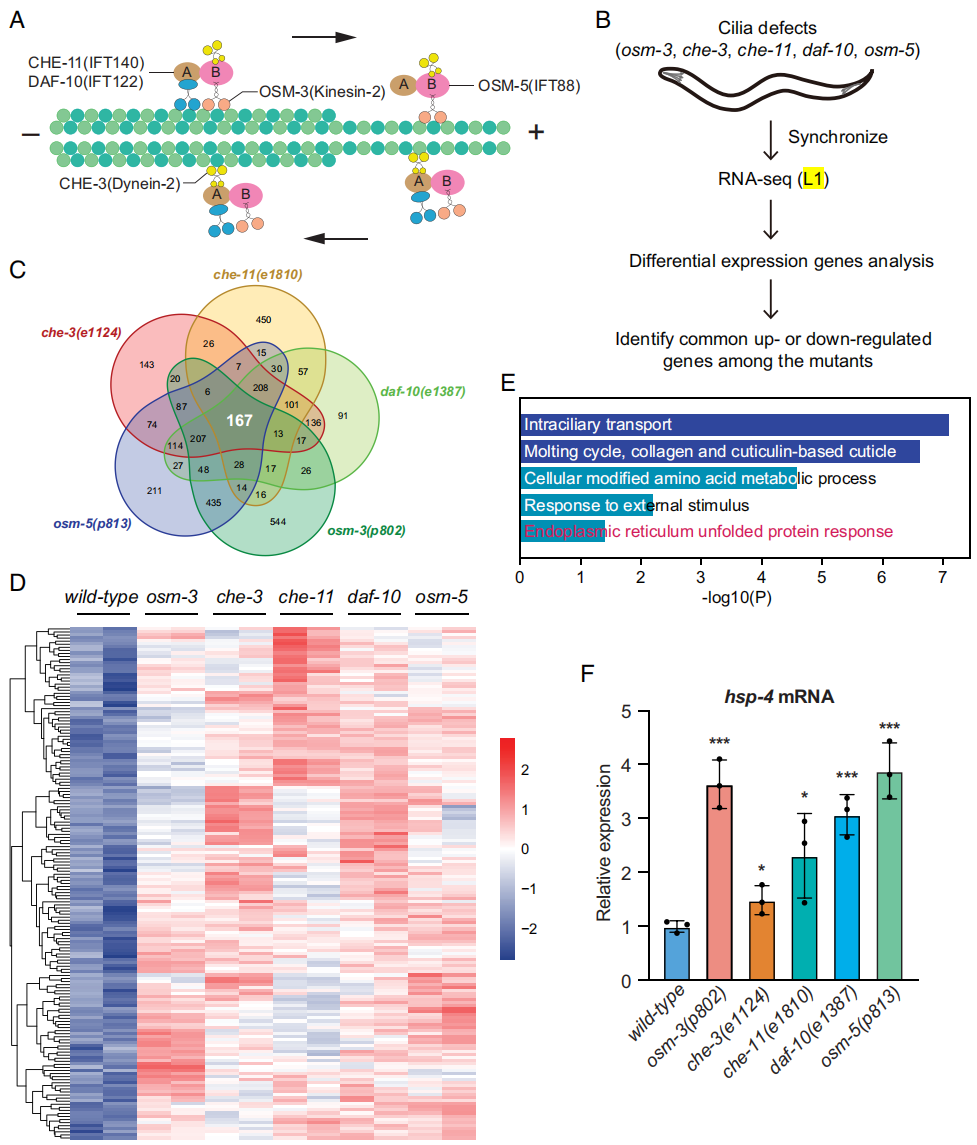
图1 纤毛突变体的RNA-seq显示hsp-4上调表达
2
纤毛应激诱导的细胞非自主UPRER是通过xbp-1分支,而非atf-6或pek-1
为探讨感觉纤毛和UPRER之间的关系,作者引入hsp-4p::GFP荧光报告基因于纤毛突变体线虫中,来检查UPRER的激活情况。结果发现野生型线虫中HSP-4的基础表达是非常低的,但是在纤毛突变体中可以观察到在远端肠道和咽部周围激活表达,这与hsp-4 mRNA表达上调的结果是一致的(图1F)。上述结果暗示纤毛应激能致远端肠道细胞产生细胞非自主UPRER。
哺乳动物UPRER不同的分支信号通路,即ATF6、PERK和IRE1α,在线虫中分别对应的是ATF-6、PEK-1和IRE-1。RNAi实验结果表明,atf-6(RNAi)和pek-1(RNAi)不能抑制hsp-4p::GFP在osm-3突变体中的表达。但是xbp-1(RNAi)则完全废除其表达,因此可认为xbp-1涉及纤毛突变体中诱导的UPRER。
线虫xbp-1的激活是通过IRE-1介导的移除其mRNA 23 nt内含子完成的,完成剪接的xbp-1最终翻译成具有活性的转录因子。文章通过将ire-1(ok799)和xbp-1(mkc62)突变体引入osm-3(p802), hsp-4p::GFP背景中,发现了ire-1和xbp-1突变体均可以抑制osm-3突变体引起的hsp-4p::GFP上调表达(图2A、B)。通过RT-qPCR衡量L4阶段线虫中被剪接xbp-1形式,即激活形式xbp-1,xbp-1s的转录本水平(图2C),结果显示除了che-3突变体外,其他纤毛突变体中均增加总的xbp-1水平(图2D)。在所有突变体中,xbp-1s表达均明显增加(图2E),且除了che-11突变体外,xbp-1s的剪接占比均增加(图2F)。另外RNA-seq IGV可视化进一步支持在突变体中总xbp-1和xbp-1s表达升高。总之,上述结果支持纤毛缺陷引起的ER应激反应依赖于IRE-1-XBP-1路径。
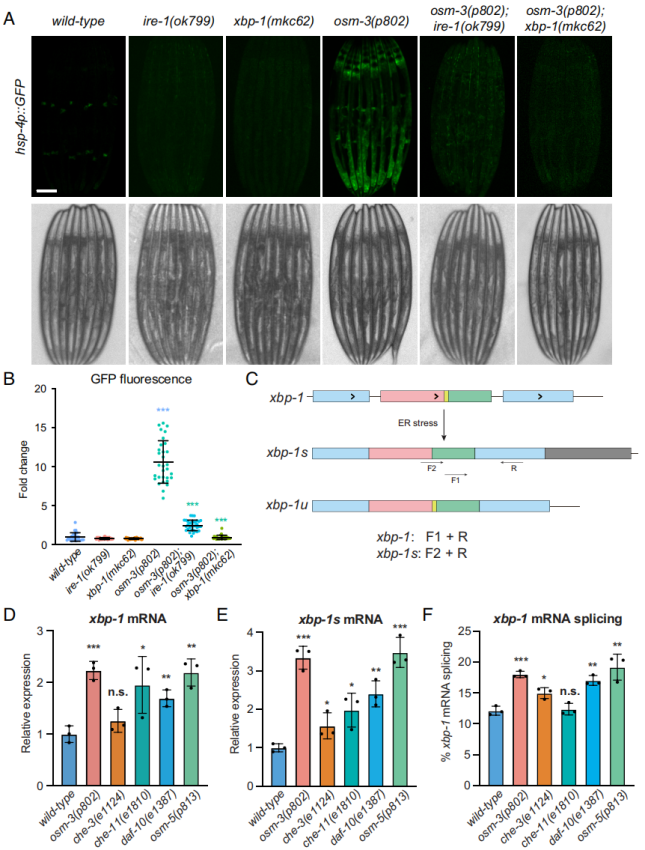
图2 纤毛应激诱导的细胞非自主UPRER是通过xbp-1分支,而非atf-6或pek-1
3
在纤毛突变体中UPRER是线虫延长寿命和衣霉素抗性所必需的
那么纤毛功能紊乱引起的UPRER激活是否涉及调节寿命的作用呢。作者发现,虽然野生型线虫与xbp-1突变体寿命没有明显差异,但是将xbp-1突变体与纤毛缺陷突变联合时,明显缩短了纤毛突变体延长寿命的表型。osm-3(p802); xbp-1(mkc62)、che-3(e1124); xbp-1(mkc62)和daf-10(e1387); xbp-1(mkc62)双突变体平均寿命达到与野生线虫相似的水平(图3A-D),表明IRE-1-XBP-1信号路径对于纤毛突变体延长寿命的表型是必需的。
衣霉素(tunicamycin)处理可抑制N乙酰氨基葡萄糖脂质中间体的形成,阻止新合成的糖蛋白糖基化,因此,在蛋白翻译活跃的细胞中可诱导形成ER应激。当野生型线虫与xbp-1突变体的L4阶段幼虫暴露于25 μg/mL衣霉素时,子代表现出幼虫发育停滞和胚胎致死的表型,而DMSO处理的对照组可自行发育至L4阶段(图3E、F)。然而,纤毛突变体展现出显著的衣霉素抗性(图3E、F)。当作者将xbp-1突变体引入这些纤毛突变体中时,它们的后代又表现出幼虫发育停滞和胚胎致死的表型(图3E、F)。转录组分析显示,衣霉素处理的纤毛突变体中xbp-1靶基因的表达显著激活。这些数据表明IRE-1-XBP-1信号路径调节纤毛突变体中UPR激活,并且对其延长寿命和衣霉素抗性是必需的。
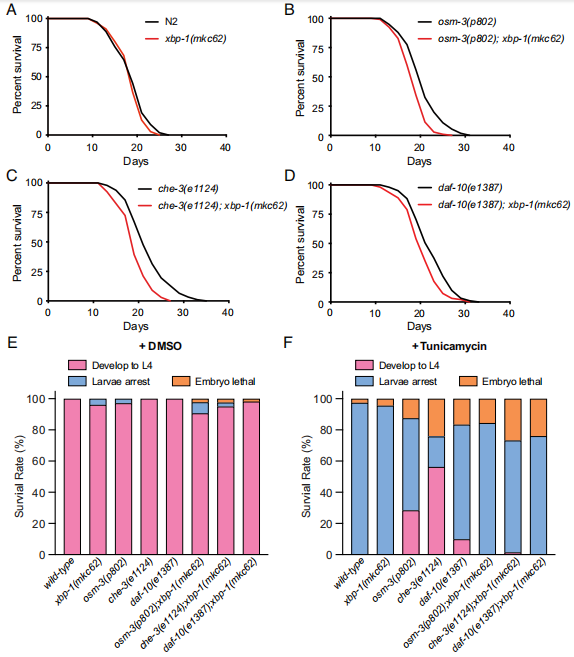
图3 在纤毛突变体中UPRER是线虫延长寿命和衣霉素抗性所必需的
4
非自主UPRER依赖于神经信号分子释放和IIS转录因子活性
文章推测末端非神经元组织中UPRER激活表明可能涉及神经内分泌信号路径,可使神经元影响到远处的UPRER状态。尽管在试验过程中osm-3和osm-5突变时均能显著上调hsp-4p::GFP表达,但是osm-5突变体展现出相较于osm-3突变体更短的纤毛结构。有趣的是,osm-3突变体线虫个体间中hsp-4p::GFP表达水平差异相较于osm-5突变体更不明显。因此,表达变化差异较小,暗示着osm-3突变体的表型更一致,所以作者在随后的试验主要关注osm-3突变体。作者以osm-3(p802); hsp-4p::GFP为背景与神经元信号分子释放缺陷型的线虫品系杂交。包括清亮小囊泡(small clear vesicles,SCV)释放缺陷线虫品系unc-13(e51)、钙离子调节的致密核心小泡(dense core vesicles,DCVs)释放缺陷线虫品系unc-31(e169)和丝氨酸类型肽链内切酶活性缺陷型线虫品系egl-3(n150)。结果显示,线虫肠道UPRER激活同时依赖于UNC-13和UNC-31功能,也与EGL-13调节的神经多肽加工过程相关(图4A、B)。
已知IIS转录因子DAF-16在纤毛突变体具有延长线虫寿命的作用。进一步探究发现,daf-16抑制是显著降低了纤毛突变体的细胞非自主UPRER激活(图4C、D)。通过比较纤毛突变体与daf-16突变体线虫的差异表达基因中发现22个上调表达和16个下调表达基因。作者同时验证WNT配体EGL-20/WNT5A在纤毛突变体中的作用,因为其涉及细胞非自主的线粒体非折叠蛋白反应(UPRMT),结果显示在egl-20突变体中UPRER没有发生改变(图4D、E)。因此,文章认为纤毛缺陷诱导的细胞非自主UPRER需要清亮小囊泡、致密核心小泡和神经多肽加工过程,同时也需要IIS转录因子。
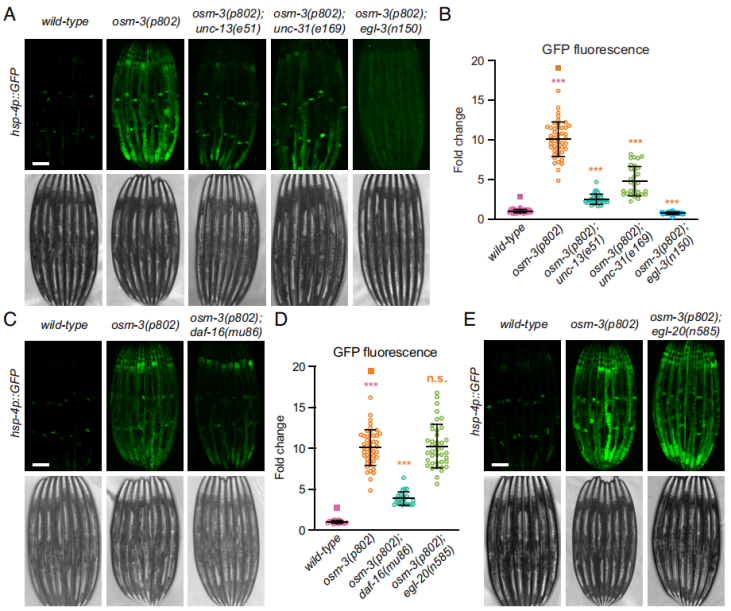
图4 非自主UPRER依赖于神经信号分子释放和IIS转录因子活性
5
压制cGMP门控通道触发UPRER和延长寿命
RNA-seq分析揭示了五个纤毛突变体较野生型有142个基因始终明显下调表达(图5A、B)。这些基因中涉及cGMP调节信号路径,暗示感觉纤毛功能紊乱与cGMP调节信号之间可能存在联系(图5C)。在线虫的感觉神经元中,cGMP的生成可激活下游cGMP门控离子通道,从而导致钠、钙离子的流入及神经元去极化。cGMP门控离子通道由TAX-4和TAX-2亚基组成,它们分别对应通道蛋白的α和β亚基。线虫TAX-4/TAX-2通道与人类CNG通道具有同源性。为了调查CNG通道是否涉及异常UPRER激活,作者将hsp-4p::GFP分别引入tax-4(p678)和tax-2(p694)突变体中。与野生型相比,tax-4和tax-2突变体明显增加末端肠道细胞中hsp-4p::GFP荧光水平(图5D、E)。重要的是,如果在tax-4和tax-2突变体中引入xbp-1突变体,则hsp-4p::GFP荧光水平与野生型相似(图5D、E)。更进一步地,在tax-4突变体中引入egl-3突变体会废除hsp-4p::GFP荧光强度升高的表型(图5D、E)。因此,压制cGMP门控通道可诱导依赖于XBP-1的UPRER。
进一步研究tax-4突变体线虫对衰老依赖性蛋白聚集的抗性,作者发现tax-4突变体在第七天降低了polyQ聚集(图5F、G),这暗示TAX-4在减少非折叠蛋白聚集中发挥重要的调节作用。
已知tax-4突变体可显著延长线虫寿命。tax-4突变体的平均寿命为29.95天,而野生型线虫为19.02天。然而在tax-4突变体中引入xbp-1突变,tax-4; xbp-1双突变体明显降低线虫寿命(图5H)。这些结果表明压制cGMP门控通道活性可能通过XBP-1调节的UPR路径以促进线虫长寿。
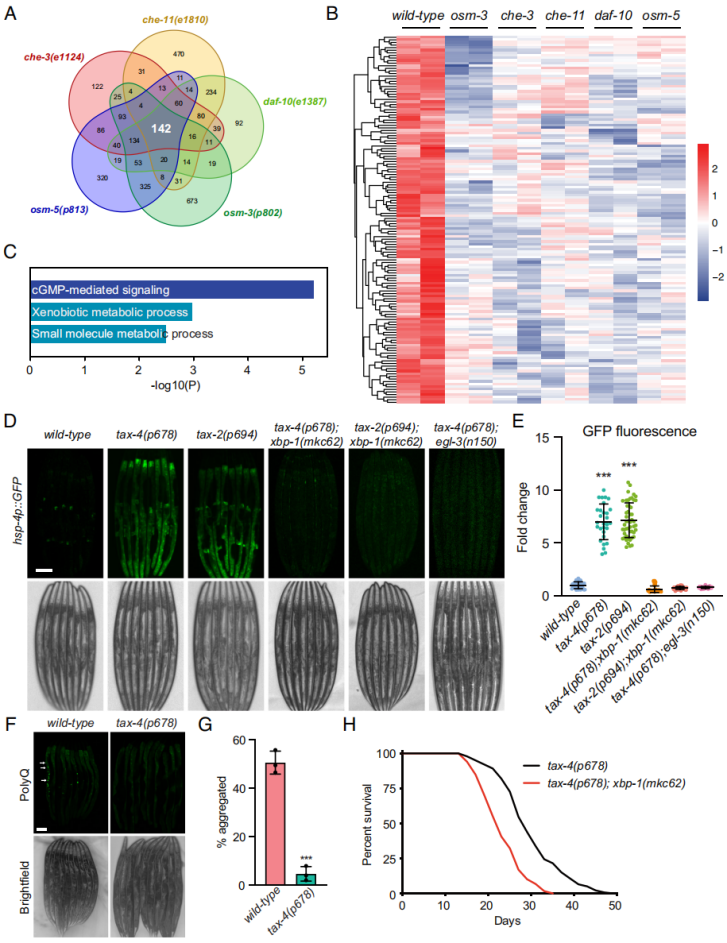
图5 压制cGMP门控通道触发UPRER和延长寿命
6
cGMP门控通道作用于纤毛的下游并激活UPRER
作者进一步提出一个问题:在纤毛突变体线虫外围组织中激活的UPRER是否是通过下游的cGMP门控通道调节的?作者检查了tax-4和tax-2突变体对感觉纤毛的影响,发现所有动物表现出正常的DiI染料摄取、纤毛长度和IFT。这暗示cGMP门控通道可能在感觉纤毛的下游发挥其功能。
对CNG通道的结构生物学和生理学研究揭示了其关键氨基酸残基的信息。在线虫TAX-4通道中第403位苯基丙氨酸突变为缬氨酸(F403V)和第407位缬氨酸突变为丙氨酸(V407A),即TAX-4(F403V; V407A)突变体,显示为TAX-4的功能缺失突变体,能够打乱cGMP结合。另外,TAX-4通道中第421的精氨酸突变成色氨酸(R421W),即TAX-4(R421W)突变体,能引起甚至cGMP不存在情况下通道组成型开放,代表一种组成型激活突变(图6A)。文章中作者在osm-3(p802); hsp-4p::GFP线虫中使用纤毛特异性的dyf-1启动子分别表达上述三种TAX-4通道蛋白。过表达TAX-4(F403V; V407A)几乎对hsp-4p::GFP没有影响(图6B、C)。但是过表达野生型TAX-4和TAX-4(R421W)显著降低hsp-4p::GFP表达(图6B、C)。重要的是,使用纤毛特异性地dyf-1启动子在tax-4(p678); hsp-4p::GFP过表达TAX-4(R421W)可减少荧光强度(图6D、E)。该结果支持tax-4神经元特异性功能,并强调其在感觉神经元中的重要作用。总之,纤毛缺陷激活的细胞非自主UPRER可能是通过降低cGMP门控通道活性来实现的。
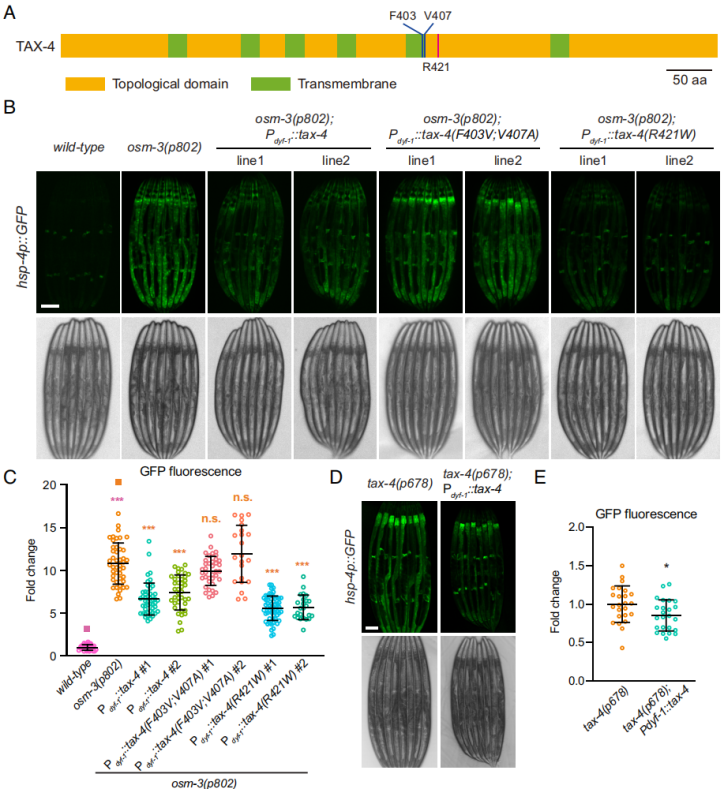
图6 cGMP门控通道作用于纤毛的下游并激活UPRER
7
化学抑制cGMP门控通道调节UPRER和寿命
人类CNGA3基因突变,例如R410W,与线虫中TAX-4(R421W)突变类似,均与细胞死亡和视锥细胞退化有关,其结果导致完全性色盲(全色盲)。已开发出小分子药物可调节CNG通道活性。其中一种化合物为ZD7288(也叫做ICI D7288)是一种有选择性的超极化激活的环核苷酸门控 (HCN) 通道阻断剂。研究评估了ZD7288对hsp-4p::GFP荧光强度的影响(图7A)。实验发现,暴露于5 mM ZD7288的线虫相较于M9缓冲液处理的对照组,其荧光强度显著增强且在线虫不同的发育阶段表现均一致,但TAX-4(R421W)过表达的线虫(Pdyf-1::tax-4(R421W))并没有影响(图7B-I)。更进一步地,ZD7288处理没有产生遗传效应,因为经处理的线虫后代在常规培养条件下,hsp-4p::GFP表达水平与对照组无显著性差异。当线虫从胚胎到成虫阶段持续暴露于ZD7288时,可延长18%的寿命。对照组最大寿命为31天,而处理组则显著增加,寿命达到了38天(图7J)。并且,过表达TAX-4(R421W)可压制长寿的表型(图7J),这进一步表明tax-4CA突变可抵消ZD7288效果的假设。这些发现表明ZD7288可延长线虫寿命。
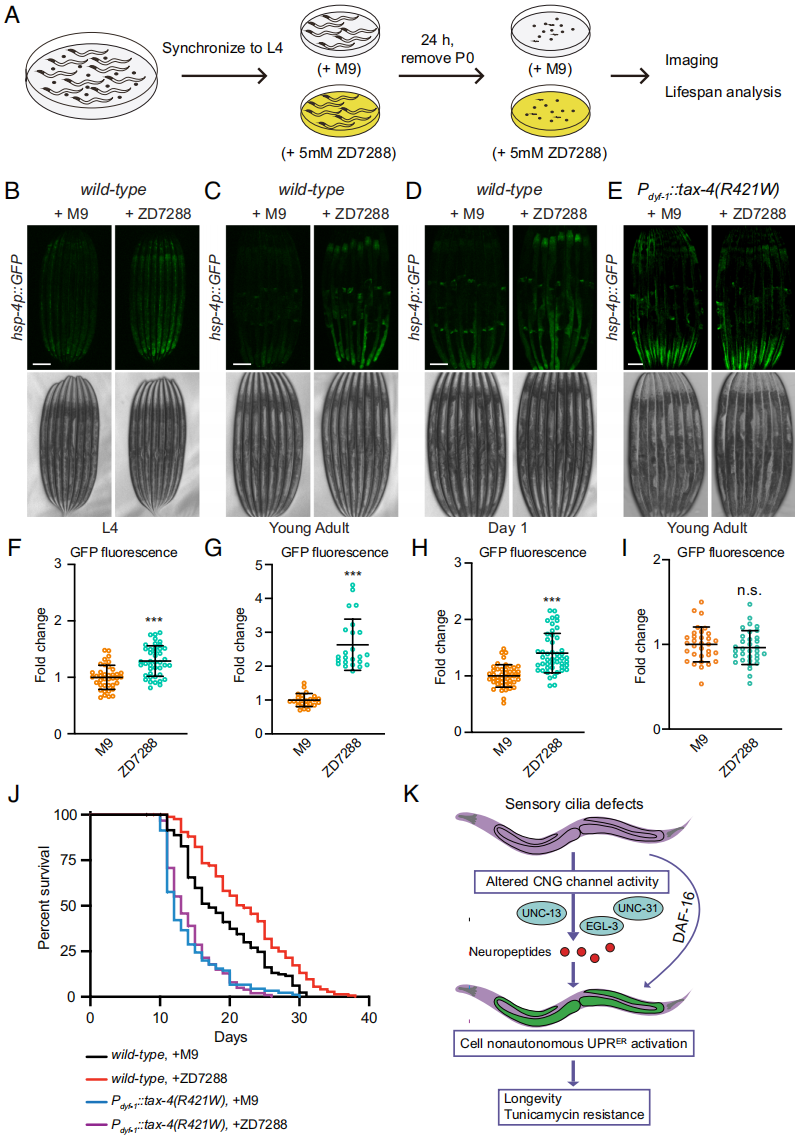
图7 化学抑制cGMP门控通道调节UPRER和寿命
8
总结
在本研究中,欧光朔研究团队发现纤毛缺陷可诱导由 IRE-1-XBP-1信号路径调节的细胞非自主UPRER,而非折叠蛋白反应对纤毛突变体产生长寿表型所必需的。并通过进一步深入研究确认CNG通道作为纤毛下游效应子触发UPRER。更重要的是文章中使用药物阻断CNG通道,可以激活UPRER并延长线虫的寿命(图7J、K)。这些发现为了解线虫感觉纤毛的生理作用提供独特的见解,并向我们展示纤毛依靠感知外部环境并调节神经活性的能力,由此引发UPRER激活最终延长寿命。而且,文章中发现通过药物策略,靶向纤毛通道,可作为潜在的促进长寿的干预方法。
撰稿:卞文印
审核:陈岚彬
编辑:余雯

